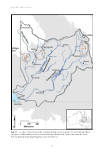
4 – Natural flows drive the ‘ boom and bust’ ecology of fish in Cooper Creek 51 First, the absolute necessity for all fish species to be able to persist in ‘refuge’ aquatic habitats (e.g. waterholes) during dry periods makes them vulnerable to any process that may accelerate rates of water loss and reduce the number of waterholes that could sustain aquatic organisms over extended dry periods. Several natural factors influence water loss by evaporation these include waterhole depth and surface area, the effective width for wind action (and evaporation), the degree of incision below levee banks, and the height and width of riparian vegetation (Hamilton et al. 2005). As waterholes decrease in depth and volume, prominent features like benches, bars, offtake channels and structural habitat (logs, riparian leaf litter and aquatic plants) become exposed and desiccated. Habitat loss reduces opportunities for fish to rest, feed and shelter from predators, leading to high mortality rates that deplete waterhole fish populations (Arthington et al. 2010). Waterholes that maintain depth, volume, habitat diversity and water quality during dry times provide refuges for fish, and their identification and protection from human interference is an essential part of fish conservation and river management during dry periods and drought (Bond et al. 2008 Silcock 2009). Human activities can influence the amount of water in waterhole refugia and the duration of waterhole persistence between flow pulses. Hamilton et al. (2005) estimated that most of the named waterholes mapped along Cooper Creek would dry within 22 months if not replenished by channel flows or flooding after 24 months of no flow, only four of the named waterholes along Cooper Creek would remain wet. However, a broader study identified over 200 waterholes in the Cooper catchment that have not dried in living memory (Silcock 2009). River flows can be altered by upstream impoundments, small-scale diversions for irrigation in the upper catchments or capture of runoff for agricultural use (water harvesting), and water can be withdrawn directly from waterholes during intervals between flows (Thoms and Sheldon 2000). Small impoundments, raised roads, or other disturbances of the geomorphological structure of anabranches can alter river flows through various channels and waterholes (Hamilton et al. 2005). Reduced flows, less frequent waterhole replenishment, and increased frequency and degree of waterhole desiccation could have deleterious ecological impacts, including reduction or local extirpation of fish and turtle numbers as well as reduced water availability for riparian trees, floodplain wildlife and livestock (Hamilton et al. 2005 Leigh et al. 2010 Sheldon et al. 2010). As well as reconnecting, refilling and replenishing isolated waterholes, channel flows can provide cues for fish spawning, movement and dispersal into backwater habitats or anabranches, allowing opportunistic discovery of high quality habitats and rich food resources or avoidance of predators. With the probable exception of Cooper Creek catfish, all members of the Cooper Creek fish community use inundated floodplains for feeding, growth, recruitment and dispersal. Hydrological connectivity between channels and floodplains is vital to these processes (Fig. 4.1). Many of the world’s floodplain rivers have lost their natural floodplain and river connections, inhibiting the natural movement patterns of aquatic species and life stages (Tockner et al. 2010). Altered catchment configurations (e.g. levee banks, roads and infrastructure) can cut off or impede delivery of flows onto floodplains (Steinfeld and Kingsford 2013), and this reduces connectivity and inhibits fish movements into favourable habitats for feeding and growth. The disruption to connectivity between
Downloaded from CSIRO with access from at 216.73.216.128 on Nov 9, 2025, 7:57 AM. (c) CSIRO Publishing